获得0积分,您同时完成了每日任务,有额外的积分奖励,请前往APP领取
立即前往
前 言
细胞因子调节干细胞增殖、自我更新及分化 | |||
因子 | 对干细胞的主要影响 | 受影响的干细胞类型 | 调节机制 |
SCF | 干细胞研究主要调节因子,与大多数细胞因子共同发挥协同或拮抗作用,细胞增殖和自我更新 | 大多数类型 | c-Kit信号通路 |
LIF | 促进细胞增殖,促进向神经元细胞分化,防止ESCs凋亡 | 大多数类型 | STAT3和MEK的激活 |
IL-1 | 加速生长 | HSCs | 转录因子Pu.1的激活 |
IL-1α | 增强骨髓MSCs的神经元发生 | NSCs、MSCs | 尚未报道 |
IL-1β | MSCs分化为成骨细胞 | MSCs | MAPK和NF-Kb的激活 |
IL-2 | 干细胞重建 | HSCs | 尚未报道 |
IL-3 | 增殖和存活 | 大多数类型 | IL-3信号通路 |
IL-4 | 在早期骨髓生成过程中非常重要,使ESCs分化为成纤维细胞样细胞,促进向心肌和骨骼肌细胞的分化 | 大多数类型 | STAT3的激活 |
IL-6 | 神经前体的调节因子,与SCF协同促进造血祖细胞扩增,对巨核系祖细胞的增殖也有重要作用,抑制骨髓MSCs向软骨细胞分化 | 大多数类型 | JAK/STAT信号通路 |
IL-7 | T细胞和B细胞的分化 | HSCs、ESCs | IL-7信号通路 |
IL-8 | 对造血和神经生成都很重要 | HSCs、NSCs | 诱导PI3K/Akt和MAPK/ERK介导的VEGF产生 |
IL-9 | 原始人红细胞的增殖,与SCF、C-Kit和其他细胞因子协同促进造血祖细胞的生长 | HSCs | STAT3的磷酸化 |
IL-10 | 增强繁殖能力,增强再生能力,c-Kit细胞中添加IL-10后获得3-4倍的HSC | HSCs | 尚未报道 |
IL-11 | 当与SCF结合时,引起小鼠造血祖细胞的扩增,对巨核系祖细胞的增殖也有重要作用 | HSCs | 尚未报道 |
IL-13 | 在早期骨髓生成过程中非常重要 | HSCs | IL-13信号通路 |
IL-15 | 从原始前体产生人类记忆干T细胞 | HSCs | IL-15信号通路 |
IL-16 | CD34+造血细胞向成熟DC的增殖和分化 | HSCs | IL-16信号通路 |
IL-27 | 直接或协同SCF诱导HSCs分化 | HSCs | IL-27信号通路 |
IL-32 | 诱导造血祖细胞增殖,降低化疗所致的骨髓毒性 | HSCs | IL-32信号通路 |
IL-33 | 促进HSCs和祖细胞的骨髓生成和髓系细胞迁移 | HSCs | 尚未报道 |
IL-34 | 对Langerhans细胞的发育必不可少 | HSCs | IL-34信号通路 |
TNF-α | 正常HSC活性的有效抑制剂,与IFN-γ协同消除MSCs的自我更新和分化 | HSCs | TNF受体p55信号通路 |
IFN-α | HSCs、NSCs的激活 | HSCs、NSCs | STAT1和AKT1磷酸化增加 |
TGF-β | 调节胚胎干细胞的异质性 | HSCs | Nodal信号通路 |
义翘神州细胞因子产品助力文章发表 | |||
货号 | 靶点 | 作用 | 文章 |
10429-HNAH | INHBA | 干细胞分化 | Shan et al. 2017 |
10008-HNAB | VEGF121 | ||
10014-HNAE | FGF2 | ||
11066-HNAB | VEGF165 | 干细胞分化 | Zluhan et al. 2016 |
10451-H08B | SCF | ||
11858-HNAE | IL3 | ||
11792-HNAH | M-CSF | ||
14890-HNAH | LIF | 干细胞培养 | Xu et al. 2022 |
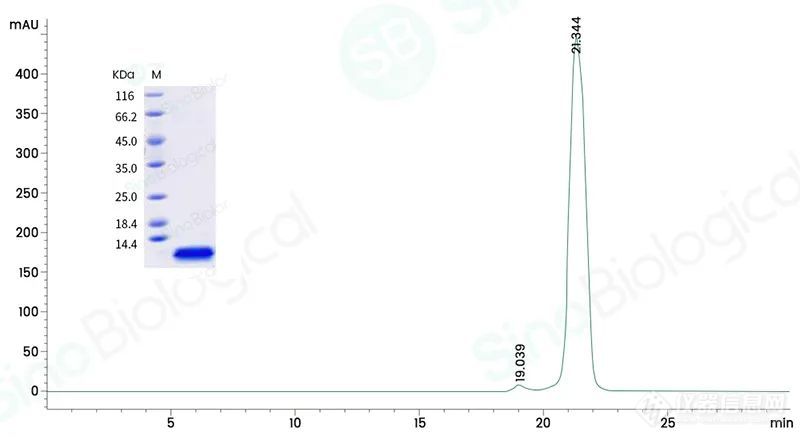
Purity: > 95 % as determined by SEC-HPLC
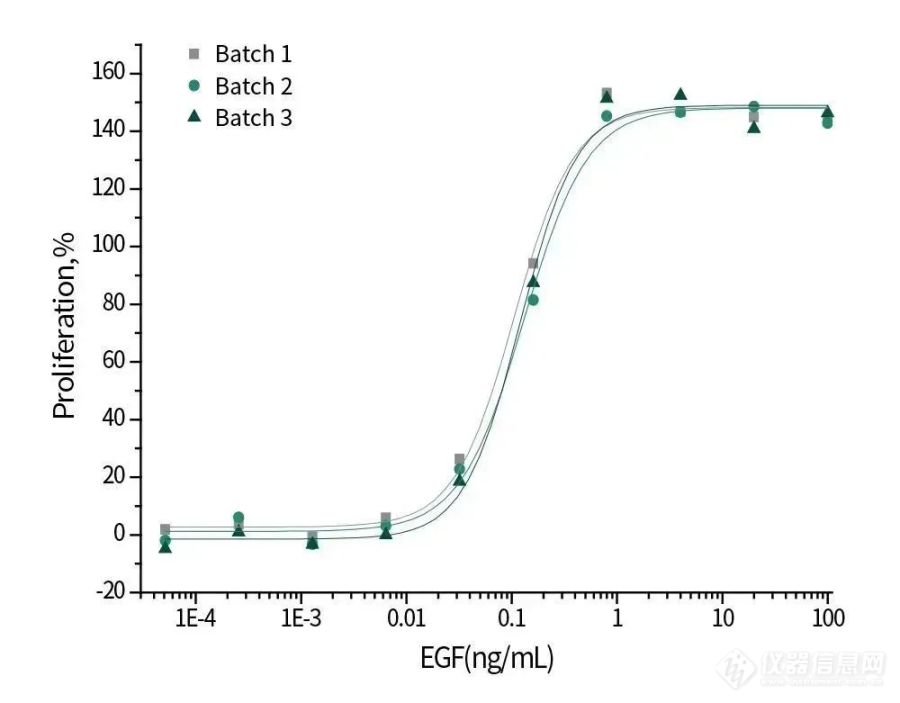
Cell proliferation assay using Balb/C 3T3 mouse embryonic fibroblasts.
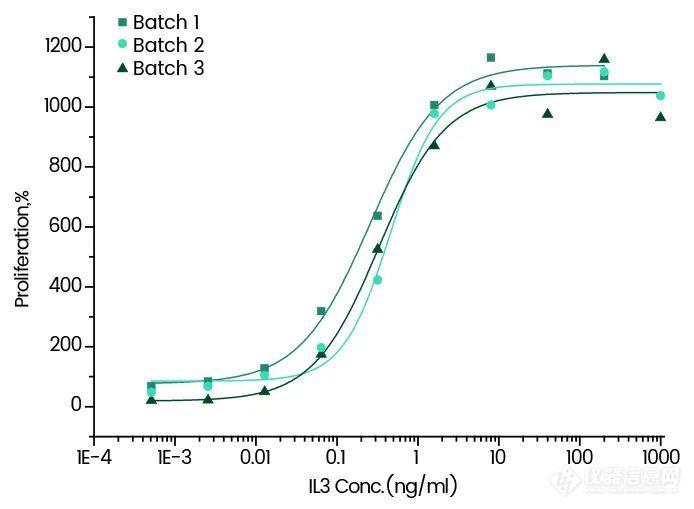
Cell proliferation assay using TF-1 human erythroleukemic cells.
更多热门人属细胞因子产品 | |||
货号 | 因子 | 内毒素 | 纯度 |
10429-HNAH | INHBA | < 10 | ≥95%☆, Active |
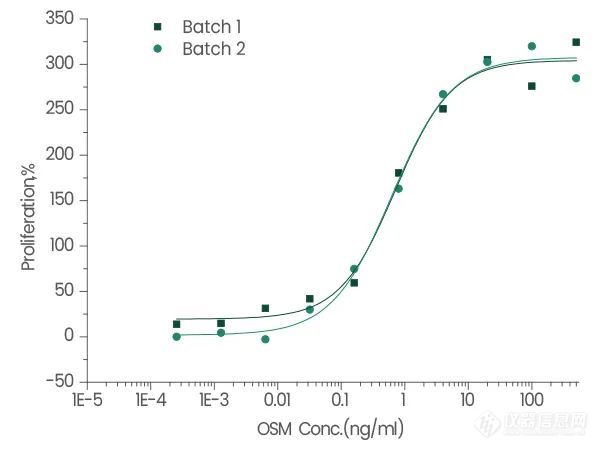
10452-HNAH | OSM | < 10 | ≥95%☆, Active |
10605-HNAE | EGF | < 5 | ≥95%☆, Active |
11066-HNAH | VEGF165 | < 10 | ≥95%☆, Active |
11858-HNAE | IL-3 | < 5 | ≥95%☆, Active |
14890-HNAH | LIF | < 10 | ≥95%☆, Active |
GMP-10014-HNAE | FGF2 | < 10 | ≥95%, Active |
GMP-10429-HNAH | INHBA | < 10 | ≥95%☆, Active |
GMP-10452-HNAH | OSM | < 5 | ≥95%☆, Active |
GMP-10605-HNAE | EGF | < 5 | ≥95%☆, Active |
GMP-11858-HNAE | IL-3 | < 5 | ≥95%☆, Active |
【参考文献】
1. W. J. Zakrzewski, et al. Stem cells: past, present, and future. Stem Cell Research & Therapy, 2019, doi: 10.1186/s13287-019-1165-5.
2. M. L. Y. Wang, Mechanism of MSCs Differentiation into Hepatocyte-Like Cells: The Role of Cytokines and Chemical Compounds. Journal of Stem Cell Research & Therapy, 2014, doi: 10.4172/2157-7633.1000179.
3. H. Li, et al. EGF/bFGF promotes survival, migration and differentiation into neurons of GFP-labeled rhesus monkey neural stem cells xenografted into the rat brain. Biochemical and Biophysical Research Communications, 2022, doi: 10.1016/j.bbrc.2022.06.077.
4. M. Mossahebi-Mohammadi, et al. FGF Signaling Pathway: A Key Regulator of Stem Cell Pluripotency. Frontiers in Cell and Developmental Biology, 2020, doi: 10.3389/fcell.2020.00079.
5. G. C. Santos et al. Leukemia Inhibitory Factor (LIF) Overexpression Increases the Angiogenic Potential of Bone Marrow Mesenchymal Stem/Stromal Cells. Frontiers in Cell and Developmental Biology, 2020, doi: 10.3389/fcell.2020.00778.